by Ananda Martins
Henry Walter Bates embarked on an incredible journey throughout the Amazon basin over 150 years ago. In addition to being amazed by the diversity of this tropical forest, he documented remarkable cases of mimicry in the vivid wing patterns of distantly related butterflies. Mimicry stands for a resemblance of different species or races, usually as the result of a selective advantage. During my PhD, I am revisiting some of the sites he explored and collecting data to test hypotheses about the evolution and spread of new wing color patterns in Heliconius butterflies, the same genus previously studied by Bates.
For my study, I focus on two species: Heliconius erato and H. melpomene. Both have some interesting biological characteristics, such as: 1) a huge diversity of colors among races within the same species, while races of different species may look very similar (Fig. 1); 2) their colorful wings warn predators of their unpalatability; and 3) specific wing color patterns are found in different regions, such that if there is a wing pattern of H. melpomene somewhere, H. erato also shows this pattern in that same region (Fig. 2).

Fig.1. Each row represents the diversity of races within species and each column the mimicry between species. First row: races of Heliconius melpomene – H. m. melpomene, H. m. thelxiope, H. m. meriana, H. m. malleti. Second row: races of Heliconius erato – H. e. erato, H. e. amalfreda, H. e, lativitta.
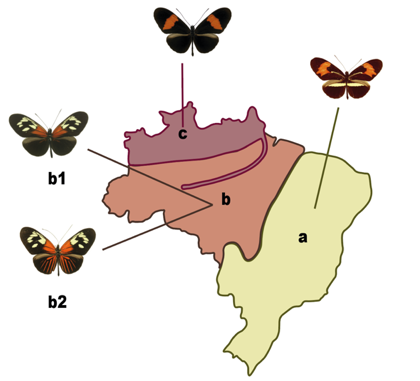
Fig.2. Geographic distribution of Heliconius wing color patterns in Brazil. Mimetic pairs of Heliconius melpomene and H. erato are found in the same regions. a. Red – yellow phenotype. b1. Dennis phenotype. b2. Dennis-ray phenotype. c. Postman phenotype.
The distribution of wing color patterns in Heliconius has long been considered the result of natural selection via predation, but other ecological factors might contribute to shaping the wing pattern distribution of these butterflies. In particular, the Amazon forest is highly heterogeneous in several environmental parameters. Thus, one of the objectives of my project is to understand which environmental factors might be contributing to the distribution of different color patterns in the Amazon. In other words, I aim to understand why we find specific Heliconius races in some regions but not others. I predict that abiotic factors are directly and indirectly selecting wing color patterns, and features like temperature, rainfall, and vegetation structure (e.g. open or closed canopy) will play important roles. For example, butterflies need to thermoregulate and differences in coloration promote different thermoregulation efficiencies. Thus, I expect that environments with higher temperatures and solar radiation will select lighter wing patterns, as lighter wings, which are less efficient at absorbing light, would not be as constrained in warmer habitats. Factors such as vegetation structure could also affect the visual signaling of Heliconius butterflies against predators or to potential mating partners, and I expect to find different Heliconius color patterns in habitats with different vegetation structure.
During the Evolution meeting of 2019 in Providence (USA), I presented some of my preliminary results. Using the Maxent software (Phillips et al., 2006) for modeling Heliconius melpomene colour pattern distributions and environmental parameters extracted from WorldClim (Hijmans et al., 2017), I found that wing color pattern distributions were shaped by environmental features, especially temperature and rainfall (Fig. 3). I also performed a cline analysis using the R package bahz (Thurman, 2019) and demonstrated that along a transect crossing different environmental gradients, as tree cover percentage, there is a gradual change in color pattern frequency (Fig. 4). This corroborates my prediction that different environments select distinct wing color patterns, and that parameters related to temperature and rainfall play a major role in races geographic distribution. Further, I suggest that other variables such as soil type and tree cover percentage might also be associated with wing pattern distribution, as proposed by Bates (1862), Brown Jr (1982), and Blum (2008). Thus, future models should include these additional ecological factors as potential predictor variables.
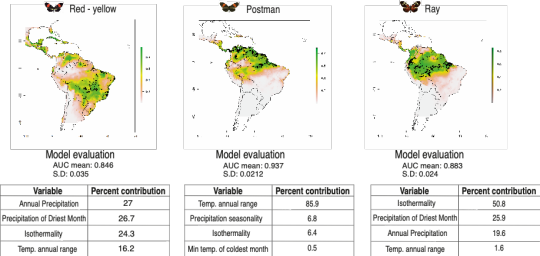
Fig.3. Geographic distribution models of red-yellow, postman and ray phenotypes of Heliconius melpomene using Maxent modeling with AUC mean model evaluation and respective standard deviation. Most important variables for each model of distribution is shown with respective percentage of contribution.
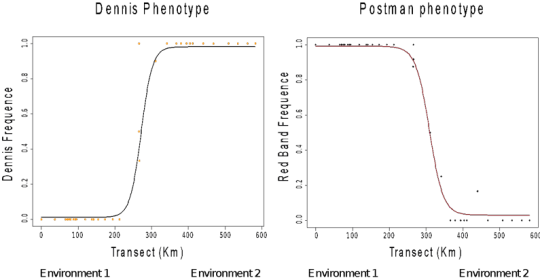
Fig.4. Cline analysis of dennis and postman phenotypes showing the change of phenotype frequency along the transect. The transect cross different environments and suggest similar selection strength for both phenotypes on opposite sides.
————————————
Em uma incrível jornada pela Amazônia há mais de 150 anos, Henry Walter Bates ficou maravilhado com a biodiversidade da floresta tropical. Além disso, ele documentou alguns casos notáveis de mimetismo (semelhança entre diferentes espécies ou raças, geralmente resultado de seleção natural) em padrões de asas de borboletas distantemente relacionadas. Durante o meu doutorado, eu estou revisitando alguns dos lugares explorados por Bates e coletando dados para testar hipóteses sobre a evolução e propagação de novos padrões de asas de borboletas do gênero Heliconius, mesmo gênero estudado por Bates.
No meu estudo foco em duas espécies: Heliconius erato e H. melpomene. Ambas apresentam algumas características biológicas interessantes como: 1) uma incrível diversidade de cores entre raças dentro da mesma espécie, enquanto raças de diferentes espécies podem apresentar padrões muito semelhantes (Fig. 1); 2) as cores vivas das asas alertam possíveis predadores que estes são animais não palatáveis; e 3) padrões específicos de asas são encontrados em regiões diferentes e, se um determinado padrão de asa de H. melpomene é encontrado em uma determinada região, o mesmo padrão de asa em H. erato é encontrado na mesma região (Fig. 2).
A distribuição de padrões de coloração de asas em Heliconius é considerada resultado da seleção natural via predação, mas outros fatores ecológicos parecem contribuir para a distribuição dessas borboletas. A floresta Amazônica, em particular, é heterogênea em diversos parâmetros ambientais. Assim, um dos objetivos do meu projeto é entender quais fatores ambientais contribuem para a distribuição de diferentes padrões de coloração na Amazônia. Em outras palavras, eu busco entender porque determinadas raças de Heliconius são encontradas em algumas regiões mas não em outras. Minha hipótese é alguns fatores abióticos estão direta e indiretamente selecionando padrões de cores das asas e características como temperatura, precipitação e estrutura da vegetação (por exemplo: dossel aberto ou fechado) apresentam papel importante nessa distribuição. Por exemplo, borboletas são animais que precisam regular a temperatura do corpo e diferenças na coloração resultam em diferenças na eficiência da termorregulação. Assim, eu espero que ambientes com maiores temperaturas e mais incidência solar irão selecionar padrões de asas mais claras, visto que asas mais claras são menos eficientes na absorção de luz e necessitam de ambientes mais quentes para uma termorregulação eficiente. Fatores como estrutura da vegetação podem influenciar os sinais visuais enviados por Heliconius contra predadores ou possíveis parceiros sexuais, assim, também espero encontrar diferentes padrões de coloração em habitats que variam quanto à estrutura da vegetação.
Durante o congresso Evolution 2019 em Providence (USA) eu apresentei alguns de meus resultados preliminares. Usando o software Maxent (Phillips et al., 2006) para modelagem de distribuição de padrões de cores em Heliconius melpomene e parâmetros ambientais extraídos do WorldClim (Hijmans et al., 2017), eu achei que o padrão de coloração de asas é moldado por fatores ambientais principalmente temperatura e precipitação (Fig. 3). Eu também realizei uma análise clinal usando o pacote do R bahz (Thurman, 2019) e nesta análise foi possível verificar que ao longo de um transecto que teoricamente está atravessando diferentes gradientes ambientais, como porcentagem de cobertura vegetal, existe uma mudança gradual na frequência dos padrões de coloração (Fig. 4). Esses resultados corroboram a hipótese de que diferentes ambientes selecionam diferentes padrões de coloração de asa e que parâmetros ligados a temperatura e precipitação desempenham um papel importante na distribuição geográfica de raças. Além disso, sugiro que outras variáveis, como o tipo de solo e a porcentagem de cobertura arbórea, também possam estar associadas à distribuição do padrão de asas, como proposto por Bates (1862), Brown Jr (1982) e Blum (2008). Assim, modelos futuros devem incluir esses fatores ecológicos adicionais como possíveis variáveis importantes.
Se você quiser continuar esta jornada comigo, siga-me no Instagram e twitter @followingbates. Estou constantemente postando notícias sobre este projeto e sobre a Amazônia brasileira.
Bates, H.W. 1862. Um Naturalista no Rio Amazonas’. 1979th edn. Edited by R. R. Junqueira. São Paulo: Universidade de São Paulo.
Blum, M.J. 2008. Ecological and genetic associations across a Heliconius hybrid zone, Journal of Evolutionary Biology, 21(1), pp. 330–341.
Brown Jr, K.S. 1982. Paleoecology and regional patterns of evolution in neotropical forest butterflies, in Prance, G.T. (ed.) Biological diversification in the tropics: proceedings, fifth international symposium of the Association for Tropical Biology. New York.
Hijmans et al., 2017. Worldclim 2: New 1-Km spatial resolution climate surfaces for global land areas, International Journal of Climatology, 37(1), pp. 4302-4315.
Phillips, S.J., Dudík, M., Schapire, R.E. Maxent software for modeling species niches and distributions (Version 3.4.1). Available from url: http://biodiversityinformatics.amnh.org/open_source/maxent/.
Thurman, 2019. bahz: Bayesian Analysis of Hybrid Zones. Available from url: https://github.com/tjthurman/BAHZ

Ananda Martins is a PhD candidate at Barrett’s Lab and associated to the Biology – Neotropical Environment program (McGill University and Smithsonian Tropical Research Institute). She is interested in patterns of biodiversity and mechanisms that drive these patterns, especially the evolution and spread of new butterfly wing color patterns in Amazon.
Instagram: https://www.instagram.com/followingbates/
Twitter: https://twitter.com/followingbates
0 Comments